大阪大学「物質・生命科学超高圧電子顕微鏡」による材料極微プロセスの時空間解析
日本電子news Vol.49 No.2
保田 英洋
大阪大学 超高圧電子顕微鏡センター、大阪大学 大学院工学研究科 マテリアル生産科学専攻
大阪大学「物質・生命科学超高圧電子顕微鏡」は、厚膜試料を高い空間分解能で観察でき、特にその立体構造をナノ~ピコメートルの尺度にまで立ち入って解明する研究や、物質内部の極微プロセスをマイクロ秒の時間分解能でその場観察する研究など、高い時間・空間分解能での材料物性の解明に威力を発揮する。本稿では、その特徴と応用研究の一例として、アモルファスアンチモンナノ粒子をとりあげ、結晶化の相転移速度やそのメカニズムについて明らかにするために、電子照射による結晶化過程をマイクロ秒時間スケールでその場観察した結果について示す。その結果に基づき、結晶核生成・成長のミクロプロセスに関して得られた知見を紹介する。
はじめに
大阪大学超高圧電子顕微鏡センターに、平成24年度国立大学法人施設整備費補助金により、世界最先端の超高圧電子顕微鏡が設置された。名称は、「物質・生命科学超高圧電子顕微鏡」である。この超高圧電子顕微鏡には、透過電子顕微鏡および走査透過電子顕微鏡としての機能がある。その最大の特長は、用いる電子線の高い透過能のために、厚さ数マイクロメートルに及ぶ厚膜試料を高い空間分解能で観察できることであり、これを活かして、デバイスから細胞にまでいたる広範な対象物の微細構造を解析する研究、特にその立体構造をナノメートルやピコメートルの尺度にまで立ち入って解明する研究に絶大な威力を発揮する。
また、この超高圧電子顕微鏡には電子直接検出型高速カメラが装備されている。これを活用することによって、物質内部の極微プロセスをマイクロ秒の時間分解能で「その場観察」できる。さらに、低温で極めて安定に稼働する試料ステージを搭載していることから、高速カメラと併用することによって、細胞など生体試料については、電子線損傷を極限まで抑えた条件下で、より天然に近い状態で観察することができる。
本稿では、物質・生命科学超高圧電子顕微鏡の特徴と、その応用研究の一例として、ナノ材料の極微プロセスをすぐれた時間・空間分解能で解明した結果について紹介する。
物質・生命科学超高圧電子顕微鏡の特徴的な性能
Fig. 1 物質・生命科学超高圧電子顕微鏡の外観

物質・生命科学超高圧電子顕微鏡(JEM-1000EES)は、Fig. 1に示すように、高さ14 m、除振装置を含む総重量330トンであり、2階にCockcroft-Walton回路の高電圧発生装置、1階に鏡体、地階に除振装置を配置した構造となっている。Fig. 2は断面図を示す。電子加速部は2つのタンクからなり、高電圧発生のためのCockcroft-Walton回路と電子加速管は、独立して収納されている。レンズ部は3段の集束レンズ、集束ミニレンズ、対物レンズ、対物ミニレンズ、 3段の中間レンズ、投射レンズからなる。Fig. 3は俯瞰図を示す。これからわかるように、超高圧電子顕微鏡本体は地下に設置された大型除振装置上に設置されている。除震装置は重量300トンのコンクリートブロックからなり、Fig. 4に示すようにそれらはエアダンパーによって浮上して支持されている。そのため、外部からの振動が電子顕微鏡本体に直接伝達することはない。超高圧電子顕微鏡の主要性能をFig. 5に示す。加速電圧は1MV、像分解能は、TEMモードにおいて点分解能0.16 nm、STEMモードにおいては1.0 nmである。像観察装置は4台のデジタルカメラからなり、モニター用、1k× 1k CCDカメラ、2k×2k CCDカメラならびに4k×4k電子直接検出カメラを装備している。
Fig. 6はシリコン結晶から撮影された高分解能像である。[110]方向から観察された格子像中には、結晶モデルに示されるように、原子間隔0.136 nmのシリコン原子が暗いコントラストによって明瞭に分離して観察されている。また、6H多形構造のシリコンカーバイド結晶の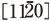 方向から観察された格子像においても、約0.11 nm間隔のシリコン原子と炭素原子を暗いコントラストで分離して観察できる。
方向から観察された格子像においても、約0.11 nm間隔のシリコン原子と炭素原子を暗いコントラストで分離して観察できる。
この超高圧電子顕微鏡の第一の特徴は、マイクロ秒時間スケールのその場観察が可能な像記録装置を装備している点である。このカメラは高感度の電子直接検出方式のデジタル画像記録が可能であり、3840×3840の有効画素数を有し、400から1600フレーム/秒の速度で画像記録が可能である。カリフォルニア工科大学のZewail博士がFig. 7に示すように、従来のビデオレートイメージングによる高空間分解能化が、将来、高速イメージングから超高速イメージングの領域へ進化すると言われている[1]。この超高圧電子顕微鏡は1フレームあたり625 μsの時間分解能を有するため、空間分解能を考慮すると赤い★印に示す超高速イメージング領域でのその場観察を可能としている。また、個々の電子を検出器で直接カウントすることにより、7680×7424の有効画素数の高精細で像撮影することも可能である。
第二の特徴は、Fig. 8に示すように、複数の試料を低温に保持できる導入機構を装備し、試料温度を100 K以下に保持して像観察を行うための対物レンズである。12個の試料を低温に保持した状態で予備室に待機させることができ、必要な試料を試料室に導入して、電子線トモグラフィー等により像観察が可能である。
Fig. 2 超高圧電子顕微鏡の断面図
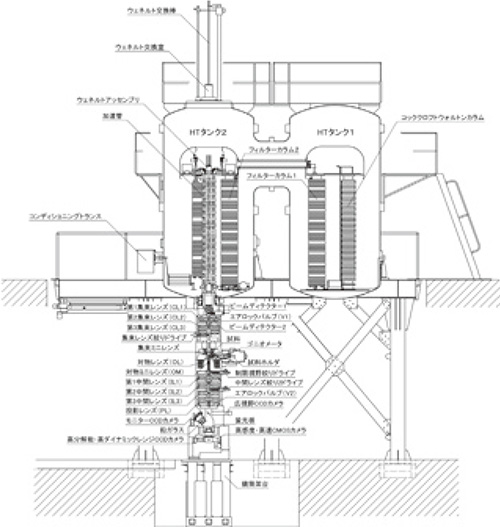
Fig. 3 超高圧電子顕微鏡の俯瞰図

Fig. 4 除振装置のエアダンパー

Fig. 5 物質・生命科学超高圧電子顕微鏡の主な性能

Fig. 6 シリコン結晶の高分解能像

Fig. 7 電子顕微鏡内その場観察における時空間分解能 [1]
![電子顕微鏡内その場観察における時空間分解能 [1]](./product_file/file/1704_07.jpg)
Fig. 8 低温試料室を備えた対物レンズ部

高速その場観察による材料研究
高速その場観察は、材料の相転移の研究に重要であり、基礎研究のみならず相転移を利用した記録材料の開発にも有用な役割を果たす[2]。従来のその場観察においては、30フレーム/秒のビデオレートイメージングが一般的であったが、最近、高時間分解能の電子顕微鏡法が開発され、材料科学において、例えば、高速の相転移、遷移状態や化学反応などの動的な現象に関する多くの応用研究が行われはじめている。中でも、パルス電子ビームを用いると、ナノ秒からフェムト秒の時間スケールでの観察が可能となる。米国ローレンス・リバーモア国立研究所においては、ナノ秒の時間分解能で電子回折のスナップショットやイメージングを用いた動的現象の観察、例えば、アモルファスゲルマニウム薄膜や金属ガラスの結晶化[3, 4]、チタンナノ結晶のα-β相転移[5]、アルミニウム薄膜の凝固[6]、Al/Ni0.91V0.09多層膜の反応による混合[7]等に、この手法が応用されてきている。
一方、連続の電子ビームを用いても、マイクロ秒の時間分解能が電子直接検出システムによって、原子スケールの分解能で実現されてきた。これは、位置敏感な電子検出器による高感度イメージングシステムと大容量の記憶装置からなる高速の記録システムである。このシステムによって、1600フレーム/秒の像記録が可能となる。
アモルファスアンチモンナノ粒子における結晶化メカニズムの解析
材料における動的な変化が重要な役割を果たす記録用アモルファス材料の結晶化の研究を、以上に示した特徴的な超高圧電子顕微鏡の性能を活用して行った。半金属のアモルファス材料は、アモルファス−結晶相転移による光の反射率、電気抵抗等の大きな差を利用した相変化型メモリー材料等に用いられ、記録容量を上げるためには記録領域を小さくする必要がある。ナノ領域における相転移の挙動はバルクとは異なるが、相転移速度やその機構は明らかではない。ここでは、光学ディスクの記録材料の成分に用いられるアンチモンに着目し、記録にはナノメートルサイズの領域が対象になることを仮定して、アモルファスアンチモンナノ粒子をとりあげる。結晶化の相転移速度やそのメカニズムについて明らかにするために、電子照射による結晶化過程を高速でその場観察し、結晶化のミクロプロセスに関して得られた知見を紹介する。
結晶化過程のその場観察
我々のグループのこれまでの研究において、アモルファスアンチモンナノ粒子は、加熱によってのみならず、外部からの刺激によっても結晶化することがわかっている。例えば、アンチモンと反応しない鉛の結晶核がアモルファスアンチモンナノ粒子の表面に接触すると、室温においても急激な結晶化が誘起されることを明らかにしている[8, 9]。一方、高エネルギー電子の照射効果を利用した相転移の研究も進めてきている。この手法を用いると、結晶からアモルファスやその逆方向の相転移が、電子のはじき出し効果により制御できる[10 ~ 12]。アモルファスアンチモン材料においては、1 MeVの高エネルギー電子により原子のはじき出しが起こるため、照射効果もアモルファスナノ粒子の結晶化のための刺激の一つになり、室温においても原子変位により結晶核が生成しうる。
以上の観点から、ここでは、室温におけるアモルファスアンチモンナノ粒子の電子照射誘起結晶化を超高圧電子顕微鏡によって評価し、結晶化の時間スケールとそのメカニズムについて議論する[13]。
アモルファスアンチモンナノ粒子はアモルファスカーボン支持膜上に蒸着法によって作製した。室温に保持したアモルファスカーボン支持膜上のアモルファスアンチモンナノ粒子を、超高圧電子顕微鏡内において加速電圧1 MV、フラックス1024 e·m-2s-1のオーダーで電子照射し、室温における結晶化過程をその場観察した。電子直接検出型カメラによって、1フレーム当たり625 μs間で原子スケールでの動画撮影を行った。
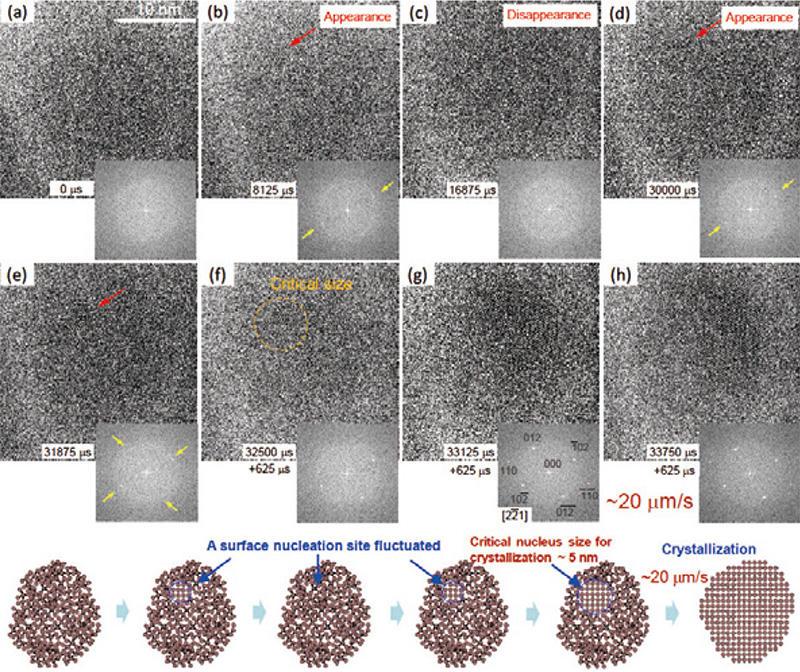
Fig. 9は、粒径約20 nmのナノ粒子の結晶化中の高分解能像のスナップショットを示す。コントラスト中のノイズの除去とアモルファスと結晶相間のコントラストを強調するためにバンドパス・フーリエ変換によるフィルター処理を行った。Fig. 9(a)に示すように、結晶化前のアモルファスナノ粒子は、ランダムなコントラストを示す。撮影に使われる電子数が少ないため、アモルファスカーボン支持膜とアモルファスナノ粒子間のコントラスト差は小さく明瞭ではない。ナノ粒子からのFFTパターン中には、アモルファス構造を示すハローリングが認められる。 Fig. 9(b)中のナノ粒子は、電子照射中の8125 μs後の同一粒子である。赤い矢印で示すように、約2 nmのサイズの結晶核が現れる。対応するFFTパターン中には、黄色い矢印で示されるように、結晶核の存在を示す2つの弱いスポットがハローリングに重畳して現れる。 Fig. 9(c)から(e)においてわかるように、ナノ粒子中の結晶核は不規則な時間間隔で消滅と出現を繰り返す。それぞれのFFTパターン中に黄色い矢印で示すように、結晶核生成による2つ、あるいは4つのスポットがしばしば観察され、それらは小さな結晶核の生成・消滅に対応している。Fig. 9(f)においては、結晶核が約5 nmまで成長し、その後Fig. 9(g)から(h)に示すように、アモルファスナノ粒子は、粒子全体にわたって結晶化する。FFTパターンにおいては、4つの弱いスポットが明瞭なネットパターンに変化し、格子定数a0=0.43 nm、c0=1.13 nmの六方晶構造のアンチモン結晶の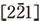 晶帯軸のパターンとして同定される。Fig. 9(f)から(h)の結果から、約20 nmのアモルファスナノ粒子において、約5 nmの結晶核が生成後の結晶−アモルファス界面の移動速度は、約20 μm s-1であると評価される。
晶帯軸のパターンとして同定される。Fig. 9(f)から(h)の結果から、約20 nmのアモルファスナノ粒子において、約5 nmの結晶核が生成後の結晶−アモルファス界面の移動速度は、約20 μm s-1であると評価される。
Fig. 9中の結晶化過程の模式図に示されているように、結晶化初期においては小さな結晶核が生成と消滅を繰り返すが、結晶核が約5 nmの臨界サイズを超えると結晶化がアモルファスナノ粒子全体にわたり急速に進行することが明らかになった。
Fig. 9 粒径約20 nmのナノ粒子の結晶化中の高分解能像のスナップショットと対応するFFTパターン
結晶成長における原子の拡散速度の評価
Fig. 10は、粒径約60 nmのナノ粒子の結晶化中の結晶−アモルファス界面の移動を示す明視野像である。それぞれのスナップショットの時間間隔は、625 μsである。Fig. 10(a)と(b)の比較から、結晶核生成サイトはナノ粒子表面であることがわかる。Fig. 10(b)から(k)において、2つの赤い矢印はアモルファスと結晶相との界面がナノ粒子の表面を交差している位置を示している。Fig. 10(b)から(f)に示すように、結晶化の初期段階においてはこの界面は小さな曲率を描くが、Fig. 10(g)から(1)に示すように、結晶化の定常状態から終了段階においては、次第に平坦になる。定常状態における界面の移動速度は、約5 μm s-1であると評価される。アモルファスナノ粒子中の結晶界面の移動を示す模式図に示されるように、結晶化はナノ粒子表面の核生成サイトから単一の結晶の連続的な成長によって起こる。Fig. 9の結果と比較すると、結晶−アモルファス界面の移動速度は粒子サイズに依存し、この観察からわかるように、粒子サイズが小さいほど界面の移動速度は速くなることが示された。
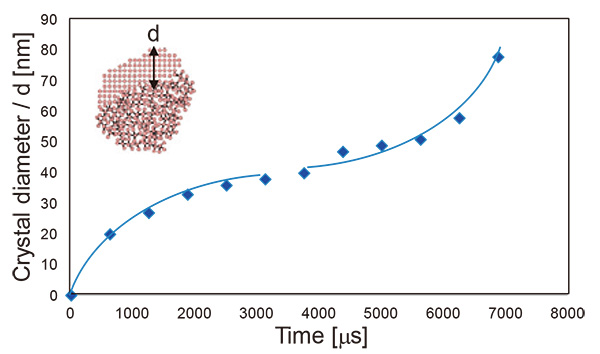
Fig. 11は、結晶核生成後の核生成サイトから界面までの距離を、時間の関数として示している。アモルファス材料において結晶核の直径dは、d=√Dt により評価される。ここで、Dは拡散係数、tは時間である。この式を用いてFig. 11の結晶成長初期のデータをフィッティングすると、Dは5.2×10-13 [m2 s-1]になると評価される。一方、バルクアンチモンの自己拡散係数DBulkは、DBulk = 2.21×10-3 exp(-2.37×104 / T)によって計算される。ここで、Tは温度である[14]。
この式を室温300 Kに外挿すると、1.08×10-37 [m2 s-1]の値が求められることから、アンチモンナノ粒子の結晶化における原子の自己拡散は、バルクにおける値に比べて少なくとも1024程度大きいことが示される。この大きな拡散係数を示す実験事実は、純物質のアモルファスナノ粒子における結晶化は長距離の原子拡散によって起こるのではなく、短距離の原子変位によって起こることを示唆している。
Fig. 10 粒径約60 nmのナノ粒子の結晶化中の結晶−アモルファス界面の移動を示す明視野像

Fig. 11 結晶核生成後における時間の関数として示された核生成サイトから界面までの距離
結晶核生成・成長メカニズム
この実験において超高圧電子顕微鏡によりはじめて達成されたマイクロ秒時間スケールのその場観察結果に基づいて、アモルファスアンチモンナノ粒子の結晶化メカニズムを議論する。
アモルファスの結晶化は、核生成と成長によって起こる。古典的な均一核生成理論によると、バルクアモルファス材料において、結晶核生成における自由エネルギー変化ΔGは、ΔG =(4πr3/3)Δ Gv + 4πr2σによって表される。ここで、rは結晶核半径、ΔGvは結晶核生成による単位体積当たりの自由エネルギーの変化(減少)、 σはアモルファス母相と結晶核間の単位面積当たりの界面エネルギー(増加)である。この古典論においては、結晶核サイズが臨界サイズよりも小さいときには、結晶核生成における自由エネルギー変化は正の値になる。その結果、結晶核は不安定になり生成と消滅を繰り返す。臨界サイズよりも結晶核のサイズが大きくなったときのみ、結晶核は安定に成長する[15]。
ここで示した実験においては、一見すると、古典的な均一結晶核生成理論によって説明されるような結晶核の生成と消滅が観察されているように思われる。しかし、ナノサイズの系における自由エネルギー変化は、結晶核生成のような局所的な構造変化に極めて敏感である。ナノ粒子中に生成した結晶核の界面近傍のアモルファス母相は、結晶界面とエピタキシャルに結晶成長しようとする傾向にある。その結果、弾性歪みがアモルファス母相中に広い範囲で誘起される。このような弾性相互作用はナノ粒子全体にわたる長距離の相互作用であり、界面の自由エネルギーを減少に導く。すなわち、こうしたナノスケールの条件下においては、局所的な急峻な界面形成に起因した系全体の自由エネルギー変化によって説明されるような古典的な均一核生成理論に基づく絶対的な臨界結晶核サイズは存在せず、臨界結晶核サイズは系の大きさによって変化する。長距離相互作用の効果により、アモルファスナノ粒子の結晶化は決められた半径rによって求められるような結晶核の自由エネルギーと界面エネルギーの単純な大小関係で核生成が起こり、その後に結晶成長が進行する2段階のメカニズムによって支配されるものではないと考えられる。この長距離の弾性相互作用がナノ粒子中の原子の協力的相互作用として働き、結晶核生成と同時に進行する結晶成長の駆動力になっていることが示唆される。
一般的にアモルファス材料において、結晶化の初期段階の核生成の潜伏期間における活性化エネルギー ΔEは、結晶成長過程のそれとほぼ等しいことから、潜伏期と成長過程が同じ原子の拡散機構によって支配されている可能性が高い[16]。特にアモルファス純物質の結晶化における原子拡散については、前述のように短距離の原子変位が支配的であることからも、核生成と成長は同時に進行すると考えられる。
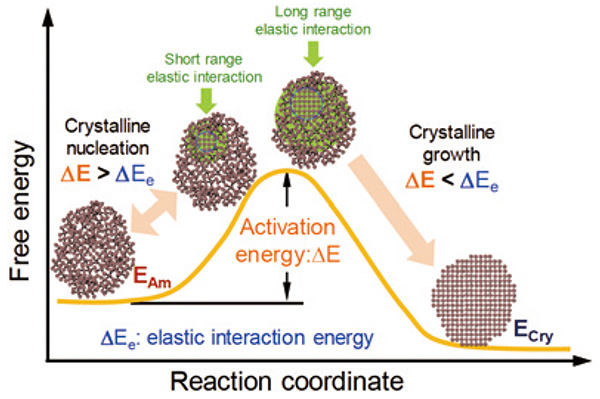
Fig. 12に結晶核生成と成長過程の模式図を、自由エネルギーと反応座標において示す。アモルファス相の自由エネルギー EAmは、結晶相のそれECryよりも大きい。結晶化の活性化エネルギーを越えるための駆動力は、ナノ粒子中の結晶−アモルファス界面近傍の長距離の弾性相互作用エネルギーΔEeによって自由エネルギーが増大することにより与えられる。図中に示されるように、短距離の原子変位によって生成した結晶核の大きさが臨界サイズよりも小さいとき、弾性相互作用は結晶核周辺に局在化するが、結晶核のサイズが増大するとともに、弾性相互作用はナノ粒子全体に影響を与える。弾性相互作用エネルギーΔEeが結晶化の活性化エネルギーΔEよりも大きくなるや否や、結晶核の成長がドミノ倒しのように短距離の原子変位によって、結晶核とエピタキシャルに進行する。
Fig. 12 自由エネルギー変化と反応座標によって示された結晶核生成と成長過程の模式図
おわりに
物質・生命科学超高圧電子顕微鏡によって、高い時間・空間分解能で物質・材料の物性の解明することが可能となった。また、低温で極めて安定に稼働する試料ステージを利用すると、細胞などの生体試料をより天然に近い状態で観察することができる。今後、こうした特徴を活かした研究を展開する予定である。
謝辞
物質・生命科学超高圧電子顕微鏡の製作において、多大なるご協力をいただきました日本電子(株)の大田繁正、大崎暁弘、我妻祐介、高桑禎將、大崎光明氏、ならびに物質・生命科学超高圧電子顕微鏡プロジェクトメンバーの皆様に感謝申し上げます。
参考文献
- A. Zewail and J. Thomas, "4D Electron Microscopy", Imperial College Press,(2010).
- D. Jacobsson, F. Panciera, J. Tersoff, M. C. Reuter, S. Lehmann, S. Hofmann, K. A. Dick and F. M. Ross, Nature, 531(2016)317.
- L. Nikolova, T. LaGrange, B. W. Reed, M. J. Stern, N. D. Browning, G. H. Campbell, J.-C. Kieffer, J. Siwick and F. Rosei1, Appl. Phys. Lett., 97(2010)203102.
- T. LaGrange, D. S. Grummon, B. W. Reed, N. D Browning, W. E. King, and G. H. Campbell, Appl. Phys. Lett., 94(2009)184101.
- T. LaGrange, M. R. Armstrong, K. Boyden, C. G. Brown, G. H. Campbell, J. D. Colvin, W. J. DeHope, A. M. Frank, D. J. Gibson, F. V. Hartemann, J. S. Kim, W. E. King, B. J. Pyke, B. W. Reed, M. D. Shirk, R. M. Shuttlesworth, B. C. Stuart, B. R. Torralva, and N. D. Browning, Appl. Phys. Lett., 89(2006)044105.
- A. Kulovits, J. M. K. Wiezorek, T. LaGrange, B. W. Reed and G. H. Campbell, Philos. Mag. Lett., 91(2011)287.
- J. S. Kim, T. LaGrange, B. W. Reed, M. L. Taheri, M. R. Armstrong, W. E. King, N. D. Browning and G. H. Campbell, Science, 321(2008)1472.
- H. Yasuda and H. Mori, Phys. Rev. Lett., 69(1992)3747.
- H. Yasuda and H. Mori, Thin Solid Films, 298(1997)143.
- G. Thomas, H. Mori, H. Fujita and R. Sinclair, Scripta Met., 16(1982)589.
- S. Anada, T. Nagase, K. Kobayashi, H. Yasuda, H. Mori, Acta Mater., 71(2014)195.
- H. Yasuda and K. Furuya, Philos. Mag. A, 80(2000)2355.
- H. Yasuda, Microscopy, 64, i27(2015).
- L. A. Gilifalco, "Atomic Migration in Crystals", Blaisdell, Waltham, MA,(1964).
- F. F. Abraham, "Homogeneous nucleation theory", Academic Press, NY,(1974).
- A. K. Varshneya, "Fundamentals of Inorganic Glasses", Academic Press,(1994).